
McCulloch wrote:Since the endogenous retrovirus is not necessary for reproduction, there is no selective pressure to keep it free from mutations, and so retroviruses will acquire mutations at about the same rate as other non-essential non-coding DNA. Eventually, retroviruses are rendered inactive because of these mutations, and they sit quietly in the genome, a testament to an infection that occurred generations in the past.
There is a contradiction in this.
“there is no selective pressure to keep it free from mutations, and so retroviruses will acquire mutations at about the same rate as other non-essential non-coding DNA.”
“and they sit quietly in the genome”
How can ERV have mutations and also sit quietly? It is either one or the other. If mutations do occur on them, then there would be significant differences between ERV in different organisms, especially if there’s a large distance of lineage between them. If mutations do not occur, then what kept them from having mutations?
Also, this assumes that endogenous retrovirus have no function. This is not the case. It has been shown in recent studies that the ERV is not functionless.
Endogenous retrovirus (ERV) elements have been shown to contribute promoter sequences that can initiate transcription of adjacent human genes.
http://www.ncbi.nlm.nih.gov/pubmed/18535086
The role of endogenous retroviruses in placental morphogenesis and trophoblast differentiation was hypothesized 10 years ago [19]. More recent studies point to the presence of HERV-R (ERV 3), HERV-FRD, HERV-W, HERV-F, HERV-K and HERV-T in human placenta, coding for intact retroviral Env proteins.
http://www.retrovirology.com/content/5/1/6
A role for endogenous retroviral sequences in the regulation of lymphocyte activation
http://www.jimmunol.org/cgi/content/abstract/143/8/2448
Thus, HERV-K (HML-2) Rec may function as an oncoprotein by de-repressing oncogenic transcription factors such as AR.
http://vir.sgmjournals.org/cgi/content/ … /91/6/1494
A better explanation of ERV is that they were originally designed in the DNA for a purpose, rather than randomly inserted into the DNA. And a prediction that follows from this is that we will continue to find more functions for ERV.
https://debatingchristianity.com/forum/viewtopic.php?p=313389#p313389
GrumpyMrGruff wrote:If my last post on ERVs was inducing ‘tl;dr’ yawns, consider the following analogy:
No, I didn’t fall asleep reading it. Actually, it was a very good post. I try to address posts in sequence, so yours was next.
GrumpyMrGruff wrote:
otseng wrote:
GrumpyMrGruff wrote: However, the authors point out that this is consistent with the current understanding of primate phylogeny. …What this demonstrates is that evolution is unfalsifiable. It can take in any evidence and present an ad hoc explanation for it.
Parsimony is key here. You’re right that Barbulescu et al. provide post-hoc explanations for the the unexpected ERV pattern. Why? Because it conflicts with the preponderance of evidence to the contrary.
If you’re referring to evidence outside of ERVs, then we can address those later. If you’re referring to ERVs, then it questions the value of it since it is open to ad hoc explanations. Also, several other assumptions of ERVs are called into question.
Is it not originally assumed that ERV is assumed to not have a function? That they just “sit quietly”?
Also, ERV was considered to be a small part of the genome, around 1%.
“In humans, endogenous retroviruses occupy about 1% of the genome, in total constituting ~30,000 different retroviruses embedded in each person’s genomic DNA.”
http://www.talkorigins.org/faqs/comdesc … troviruses
However, the number discovered is now closer to 8%.
“There are many thousands of endogenous retroviruses within human DNA (HERVs comprise nearly 8% of the human genome, with 98,000 elements and fragments[9]). All appear to be defective, containing nonsense mutations or major deletions, and cannot produce infectious virus particles.”
http://en.wikipedia.org/wiki/Endogenous … troviruses
How did 8% of the genome originate from mistakes from viruses? This is more problematic if most of these are found to have a function.
Also, the prediction that I mentioned before is that we will continue to find functions for ERV and that they will not all just be considered “defective” and “containing nonsense mutations”. However, if they are all indeed found to be completely functionless, it would make more sense that it is an inactive remnant from an ancestor.
Stating this more strongly, comparative genomics would falsify the concept of species descent/modification if resampling different traits (different genes, ERVs, etc.) inferred the same tree no more frequently than when using randomized data. In other words, all noise and no signal would be evidence against shared ancestry.
If ERV is indeed functionless, yes, what you stated would make sense.
The preponderance of genomic sequence data (including ERVs), mtDNA sequence data, karyotypic evidence, and fossil distribution are consistent with a single ancestral tree.
However, as for actual fossil evidence of a common ancestor, it is lacking. As for genetic similarities, if species share morphological similarities, it would make sense that they also share genetic similarities.
You should be careful with your phrasing, though. You ask Goat about “an ERV … found in humans and not in chimps” – there are many such ERVs. However, they’re also absent in gorillas, orangs, and other primates… because we have acquired them after our divergence from chimps (and all other primates).
I do not believe I said that. I did state: “How about if I find an ERV common to primates (including chimps), but not found in humans?”
Incidentally, this seems at odds with your earlier claim that the host-specific functions of some ERVs imply their design. Different human populations have different total numbers of ERVs. While we all have some ERVs in common (including those inherited from our common ancestor), various reproductively isolated human populations have accumulated different ERVs at different positions in their genomes. Just as inter-species ERV distributions make sense in light of evolution, this pattern makes sense in light of virology and population genetics… but not in light of your assertion that ERVs were specifically engineered in the genome. If they’re designed to serve a purpose, why do some human populations need different types and numbers of ERVs? Did the Designer continue to tinker after Adam and Eve?
If there are ERV differences in humans, it would not show that humans evolved. It would only either show that a virus infected a certain group or that there would be differences in genotype/phenotype in populations.
https://debatingchristianity.com/forum/viewtopic.php?p=315590#p315590
GrumpyMrGruff wrote: This characterization only holds with respect to viral behavior. As has been previously pointed out, spawning actively infectious retroviruses isn’t good for survival. However, loss of viral activity has no bearing on whether natural selection jury-rigs an ERV for host-beneficial functions after it is in the host genome. Conversely, some ERVs contain viral genes which are not completely inactivated and can promote retrovirus-related diseases such as cancer[1] and MS.[2]
Which leads me to wonder, is there data showing roughly what percentage of mutations result in detrimental, neutral, and beneficial results?
And another question, how do geneticists decide if something is an ERV?
1% versus 8% is a bit of a non sequitur. More sequence information was processed between 2000 (1%) and 2004 (8%), leading to more identified ERV sequences.
It seems peculiar that if an ERV insertion would be a remote event, that 8% of our entire genome would consist of leftover virus injections.
You’re also focusing on a false dichotomy here. There is no reason to assume that ERVs must be completely inactive genetic elements OR have host-beneficial functions.
It seems to me there would be three possibilities of viral injections – pathogenic, neutral, functional. If it was pathogenic, it would cause the destruction of the cell, so the only thing left would be the other two.
Also, the prediction that I mentioned before is that we will continue to find functions for ERV and that they will not all just be considered “defective” and “containing nonsense mutations”. However, if they are all indeed found to be completely functionless, it would make more sense that it is an inactive remnant from an ancestor.See the above regarding function. Again, why is it either-or? Why do they all need to be completely functionless?
It doesn’t. My point is that if all ERV are found to be defective, then the most reasonable explanation is a common ancestor and not a common designer. If more functions are found, it becomes less reasonable. If a large majority of ERV are to be found with function, then it would be more reasonable to accept that it was designed.
I’ll make my own prediction, though it may be a while before I can dig up the relevant papers: ERV sequences shared by all primates, all mammals, etc. (and therefore thought to have been acquired by a distant ancestor) will be more likely (on average) to have host functions than ERVs shared by only great apes or those found only in humans (or recently acquired ERVs in any other genus/species). Why: Mutation and natural selection will have had more time to co-opt old ERVs than those incorporated only recently.
Makes sense, I accept your prediction.
This line of reasoning might hold in genes and regulatory elements controlling morphology (e.g., homeobox genes, bone morphogenetic proteins, hedgehog genes, etc., and their promoters), but it cannot logically be extended from morphological features to all genetic sequences in an organism.
Yes, I agree that it cannot be extended to all genetic sequences.
Bear in mind that regardless of whether an ERV has acquired a host-beneficial function, it will most likely have its own pathogenic genes broken by mutation (a few known examples don’t – cancer, MS, causing ERVs).
Another question. Is it possible to compare an ERV and a virus genome and show that an ERV was derived from an actual virus? Can it be determined what actually mutated?
Conversely, convergent evolution of all these inactive ERVs from different initial states is exceedingly unlikely.
I would agree. And this can be ruled out.
https://debatingchristianity.com/forum/viewtopic.php?p=316062#p316062
GrumpyMrGruff wrote:
otseng wrote:Which leads me to wonder, is there data showing roughly what percentage of mutations result in detrimental, neutral, and beneficial results?Sorry, when you say mutations do you mean ERV insertions or any mutations? In the former case, I don’t know of any papers trying to quantify the ratio of outcomes. In the latter case, there are many papers about the outcomes of single-nucleotide mutations in genes (whether silent (neutral) or not, and whether non-silent mutants are detrimental (many) or beneficial (a few)).
First let me say that I appreciate your thoughtful responses and patience in answering my questions.
When I read posts, I try to understand to the best of my ability what the poster is saying. And since this is an area I’ve never studied before, it’s taking me awhile to research and understand the concepts.
OK, back to answering your question.
Either case (ERV mutations or all mutations) would be fine. Just curious about what is roughly the percentage values.
And another question, how do geneticists decide if something is an ERV?During the part of their life cycles when retroviruses insert themselves into host genomes, they look like this.
Figure 1: Schematic of retroviral genome arrangement
Figure 3: Schematic of retrotransposon structure
Look familiar? The POL gene (under the control of the LTR) allows the retrotransposon to copy itself. Without working copies of the capsid proteins (GAG) or envelope proteins (ENV), it cannot make new virus particles.

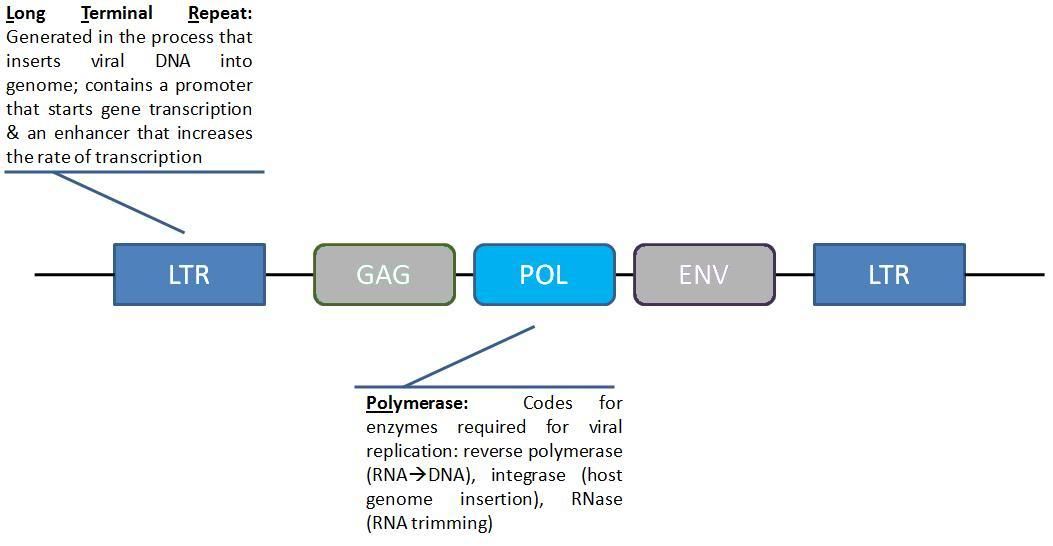
Since ERV and retrotransposon are of similar structure (seemingly identical to me), does that mean that all retrotransposons are an ERV (or can trace its source to an ERV)? If not, how can one distinguish between the two?
To find ERVs, researchers look at genomic sequence data for retroviral genes. Possible hits may look like these (many others are possible, but you get the idea):
Figure 2: Some possible ERV structuresThe first case is the equivalent of a “completely intact fossil.” All of the genes found in live retroviruses are present (though inactive) and in an order corresponding to a living class of retrovirus.
The second case gives another example of a “completely intact fossil,” albeit a partially active one. The LTR still promotes expression of the GAG gene (it could easily be another viral gene), though without the full suite of retroviral genes this ERV is not pathogenic (in most cases). Sometimes expression of partially active ERVs can lead to autoimmune response[1]. Conversely, these genes (though initially neutral) may be co-opted by natural selection to serve host-beneficial functions.[2]
The remaining classes are “partial fossils.” These partially deleted fragments may or may not have regulatory activity or gene expression, but they match the general arrangements of other retroviruses.

I understand the first and second cases, but for the others, is it simply because of the existence of a LTR that shows it originally was an ERV and has experienced deletion of material?
Granted, but there’s nothing to stop a neutral ERV from gaining a beneficial function. Regardless of whether such a beneficial mutation is passed to subsequent species, creationists shouldn’t take issue with the initial neutral-to-beneficial mutation – it would be an example of “microevolution” within a species.
I do not totally discount the possibility of a mutation to result in a beneficial function. But my guess is that the percentage of such mutations would be significantly less than detrimental mutatations.
Far less than 1% of known ERVs show this function – didn’t the creationist position hinge on finding many, many functional ERVs?
Since transposons were only discovered relatively recently, I think it would be fair to say that there is still much research that can be done in this area. If further research confirms that ERVs have no function, I acknowledge that it would be more plausible to explain it by evolutionary common descent.
Far less than 1% of known ERVs show this function – didn’t the creationist position hinge on finding many, many functional ERVs?
It would be a prediction, yes, that a significant portion of ERVs should have function. And if future research confirms that ERVs are functionless, it would falsify this prediction.
Why do we observe inactive viral genes that are not required for the function of syncytin? Why does a designer include these functionless components when the ENV/syncytin is the only part that’s required?
If we do not know the function of something now, it does not necessarily mean that it is actually functionless.
Take for example introns:
Introns may also contain “old code”, or sections of a gene that were once translated into a protein, but have since become inactive. It was generally assumed that the sequence of any given intron is junk DNA with no biological function. More recently, however, this is being disputed.
http://en.wikipedia.org/wiki/Intron
And even all ERVs were also assumed to be functionless, but research is starting to reverse this assumption.
Since LTRs are all that is required for gene regulation, why do we often see inactivated viral genes along side them (Figure 5)? These genes may be mutationally reactivated to cause diseases like cancer or MS.
When you say “mutationally reactivated”, it would imply the purpose of the original sequence was to cause cancer or MS.
The most parsimonious explanation for these viral relics is retroviral insertion and subsequent natural selection. And if retroviral insertion is truly the cause of ERVs (functional or not), then the incredible odds against random insertion of the same viral strain at the same genomic locus across species provides incredibly strong evidence for shared ancestry. (The inescapable fact creationists were trying to avoid by invoking design of ERVs.)
Again, I would agree with this if ERVs are actually functionless. But, would you also agree that if a signficant portion of ERVs are found to have function, then the better explanation is purposeful design?
https://debatingchristianity.com/forum/viewtopic.php?p=317735#p317735
GrumpyMrGruff wrote:At no point in my replies am I using the word ‘function’ in a teleological sense.
I understand. And I’m not trying to infer in any way that you subscribe to any teleological cause.
We have observed speciation via natural selection in the past. Your best defense is to demonstrate biological mechanism(s) which would definitively prevent the divergence of primates into humans, chimps, and gorillas.
Yes, I would agree that we have observed speciation. But, it would be quite an extrapolation to show that this demonstrates evolution of (non-human) primates into humans. So, rather than placing the burden on me to disprove this, the burden is on those who claimed that this indeed has happened.
‘I can’t believe it happened’ is often deployed as an argument from incredulity in creationist circles, but I’ve never seen them present the mechanism(s) for it.
If you mean a mechanism for microevolution, I have no disagreements with it. If you mean an alternative explanation for the origin of man, I’ve presented the human creation model.
Again, I would agree with this if ERVs are actually functionless. But, would you also agree that if a significant portion of ERVs are found to have function, then the better explanation is purposeful design?I assume that you are using function in a teleological sense.
No. I use “function” to mean that it performs a specific beneficial task. The function of the heart is to pump blood. The function of the eye is to see. And I’m not stating that just because something has a function that it was designed by a designer.
So to answer your question: No, if a significant portion of ERVs are found to have host-beneficial behavior, I would still have no way of inferring that they were purposefully designed.
I take this as all evidence for ERVs, no matter if they are found to have function or not, would always be evidence for evolution.
Until an observable species designer (kinds designer?) is produced, the design scenario remains less parsimonious than evolution because it relies on mechanisms we haven’t observed.
If we are to only go by what has been observed, then we can also rule out macroevolution. Macroevolution would only be an extrapolation of microevolution, not something that we can observe.
However, there are other retrotransposons that behave similarly (copy/paste) but are structurally very different.
I realize that the diagrams you presented are highly simplified, but at the risk of getting too technical here, how can one distinguish between an ERV retrotransposon and a non-ERV retrotransposon?
Deletions can remove LTRs (see the last example in the figure).
It would appear that deleted sequences would be common in the genome as evidenced by left-over ERVs. Would deletions be an entirely random event? What would cause it?
Don’t fall into the same conceptual trap that many creationists do. I too suspect that most mutations are either harmful or neutral. But natural selection (a repeatably observed and uncontested mechanism in biology) acts as a filter. Hence we expect organisms to accumulate the relatively rare beneficial mutations over generations, while harmful mutations remain at relatively low levels in populations.
I think I’ve already fallen into that trap. ![]()
However, it would not account for neutral mutations. And also, I’m not convinced that harmful mutations are filtered out in humans.
Again, how can their host-beneficial behavior allow inference of design? (Not at all a rhetorical question!)
The reasons I would say that if a significant portion of ERVs are found to have beneficial functions would be an indicator of intentional design are two-fold. One is that neutral mutations would not be affected by natural selection, so there would be no mechanism to select them out. Another is that I do not see any evidence that harmful genetic mutations in humans are actually filtered out by any natural selection process. Take for example sickle cell disease. There is no indication that it will eventually disappear by natural selection or any other genetically inherited diseases.
https://debatingchristianity.com/forum/viewtopic.php?p=319796#p319796